 Intranet
Intranet Access
Access Contact
Contact
Bacterial regulatory RNAs: function, mechanisms, regulatory networks
We exploit the genomic data obtained from Staphylococcus aureus in order to establish function of regulatory RNAs, their structures, their modes of actions, and their regulatory networks to which they belong. High-throughput technologies (as quantitative proteomic and RNAseq) are combined with molecular and structural biology tools. In particular, we explore the role of RNAs in bacterial physiology (metabolism, stress response), in cell-to-cell communication within a monoclonal (quorum sensing system) or heterogeneous (biofilm) population, in virulence and pathogen-host interactions.
Regulatory RNAs in Staphylococcus aureus
Investigator: Dr. Isabelle Caldelari
Four major axes are developed:
- Identification of targets and mode of action of RNAIII, the intracellular effector of quorum sensing
- Function of novel non-coding RNAs and their regulatory networks
- Analysis of regulatory mechanisms involving 5’ or 3’ untranslated regions of mRNAs
- Development of experimental strategies to characterize direct targets of RNAs and their protein machinery
S. aureus produces several virulence factors, whose synthesis is controlled temporally by two component systems, transcriptional factors and regulatory RNAs. The agr system discovered by R. Novick in 1993 senses cell density; it encodes two intracellular effectors, the response regulator AgrA and RNAIII. RNAIII is a multifunctional RNA, coding for delta haemolysin and acting as an antisense RNA to regulate translation and degradation of RNA targets. Using a similar mechanism, we showed that RNAIII inhibited the synthesis of adhesion factors, the transcriptional repressor of toxin Rot, and of enzymes involved in peptidoglycan metabolism as LytM. By repressing Rot, RNAIII activated indirectly the synthesis of exotoxins. This double feedforward loop motif behaves as a selector switch allowing the bacterial population to disseminate and colonise tissues at high cell density.
We have contributed to the identification of more than 50 non-coding RNAs (ncRNA). A class of ncRNAs contained a conserved motif rich in cytosines, as in RNAIII, which interacts with the Shine and Dalgarno sequence of repressed target mRNAs. As examples, RsaE inhibited the synthesis of enzymes of the TCA cycle and regulated the central metabolism at the beginning of the stationary phase of growth. The two component system SrrAB, which senses anaerobiosis and the presence of free radicals like NO induces the transcription of RsaE. A second example is RsaA whose expression is induced by the sigma B factor. RsaA represses the synthesis of the pleiotropic regulator MgrA. Subsequently RsaA activates biofilm formation, represses the synthesis of capsule and favors opsonophagocytosis. Our data illustrate intricate relationships among regulatory networks to integrate various signals in addition to quorum sensing to control virulence.
We aim to characterise functional interactions between RNAs and transcriptional factors and to follow the networks dynamic in the bacterial population.
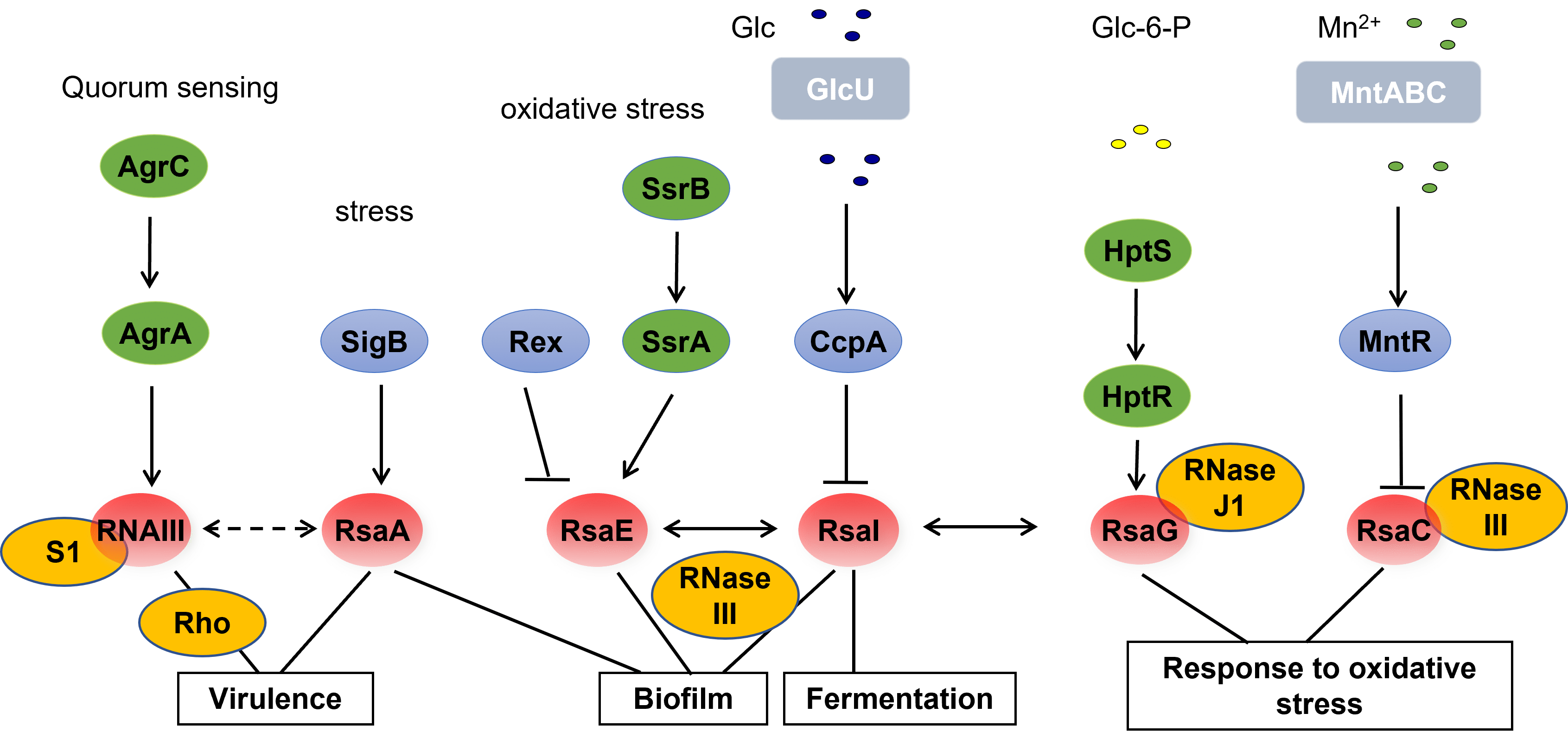
The endoribonuclease III (RNase III) cuts double-stranded RNAs and is one of the enzymes associated with antisense RNA regulation. Using RIP-Seq, we identified numerous RNA targets and revealed new mechanisms involving RNase III: maturation of mRNA leading to a better stabilisation and translation, maturation of divergent or convergent mRNAs possessing complementary untranslated regions, and regulation of pervasive transcription.
What is the impact of the co-maturation of divergent or convergent genes on the physiology of the bacteria? Are other enzymes or proteins playing a role in RNa-dependent regulation?
To map the ncRNA targetomes (identification of direct RNA targets) and helper proteins involved in post-transcriptional regulation, we applied to S. aureusseveral approaches in vivo derived from literature. These methods are based on separation of the complexes either by glycerol gradient or using affinity chromatography with tagged mRNA/ncRNA. We then identify RNA targets and protein ligands with RNAseq and mass spectrometry, respectively.
These approaches establish RNA-RNA interactome and the deduced regulatory networks. Functions and mode of actions of several RNA co-factors are actually studied.
