 Intranet
Intranet Acces
Acces Contact
Contact
ARN régulateurs bactériens : fonction, mécanisme, réseaux de régulation
Nous exploitons nos données génomiques obtenues sur Staphylococcus aureus pour établir les fonctions des ARN régulateurs, leur structure, leurs mécanismes d’action et leurs réseaux de régulation. Pour ce faire, nous utilisons des technologies de pointe (approches haut débit en protéomique et RNA-seq) combinée à des méthodes plus classiques de biologie moléculaire, génétique et structurale. Nous explorons en particulier, l’implication des ARN dans la physiologie de la bactérie (métabolisme, réponse au stress), dans la communication entre cellules au sein d’une population monoclonale (système de densité cellulaire) et hétérogène (biofilm), dans la virulence, et dans les interactions hôte-bactérie.
Les ARN régulateurs de Staphylococcus aureus
Responsable scientifique: Dr. Isabelle Caldelari
Quatre axes majeurs sont développés :
- Identification des cibles et mécanismes d’action de l’ARNIII, effecteur intracellulaire du système de densité cellulaire
- Fonction de nouveaux ARN non codants et leurs réseaux de régulation
- Analyse de mécanisme de régulation impliquant les régions 5’ et 3’ des ARNm
- Mise au point de stratégies expérimentales pour caractériser les cibles directes des ARN et les protéines associées à leurs fonctions
S. aureus produit de nombreux facteurs de virulence dont la synthèse est contrôlée de manière temporelle par divers régulateurs incluant des systèmes à deux composants, des régulateurs transcriptionnels et des ARN. Le système agr découvert par R. Novick en 1993 répond à la densité cellulaire ; il code pour deux effecteurs intracellulaires, le régulateur de réponse AgrA et l’ARNIII. L’ARNIII est un ARN multifonctionnel qui code pour l’hémolysine delta alors que les régions non traduites agissent comme antisens pour réguler la traduction et la dégradation des ARNm cibles. Nous avons en particulier montré que cet ARN réprime par un mécanisme commun la synthèse de facteurs d’adhésion et du facteur Rot répresseur de la transcription des gènes codant les exotoxines ainsi que des enzymes impliqués dans le métabolisme du peptidoglycane comme LytM. En régulant le facteur de transcription Rot, l’ARNIII active indirectement les exotoxines et permet aux bactéries de se disséminer et de coloniser d’autres tissus à haute densité cellulaire.
Nous avons contribué à l’identification de plus de 50 ARN non codants (ARNnc) dont plusieurs d’entre eux possèdent un motif conservé riche en cytosines aussi présent dans l’ARNIII, qui serait un site d’amorçage pour interagir avec la séquence Shine et Dalgarno des ARNm cibles réprimés. Ainsi, RsaE réprime la synthèse de plusieurs enzymes impliqués dans le cycle TCA et régule le métabolisme central à l’entrée de la phase stationnaire. La transcription de cet ARN est induite par le système à deux composants SrrAB qui répond à l’anaérobiose et à la production des radicaux libres NO. RsaA, dont l’expression est induite par le facteur Sigma B, réprime la synthèse du régulateur pléiotropique MgrA, et en conséquence induit la formation du biofilm, réprime la synthèse de la capsule et favorise l’opsonophagocytose de la bactérie. RsaA est au cœur d’un réseau de régulation qui favorise l’installation d’une infection chronique. Ces résultats illustrent la complexité des interactions entre réseaux de régulation permettant d’intégrer d’autres signaux autres que la densité cellulaire pour réguler les facteurs de virulence.
Nos objectifs sont de caractériser les interactions fonctionnelles entre les ARN et d’autres régulateurs de la transcription et de suivre la dynamique de ces réseaux au sein de la population.
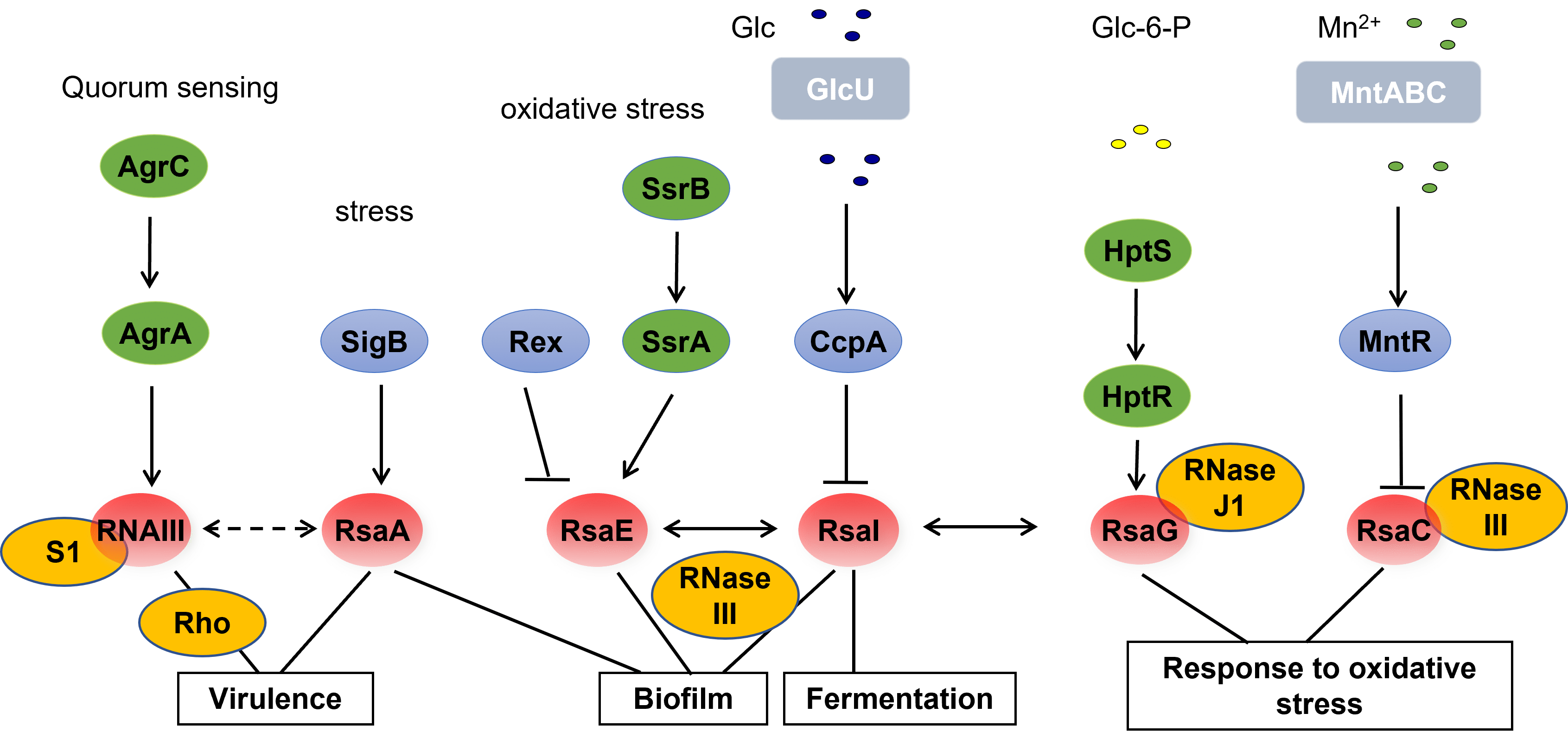
L’endoribonucléase III qui coupe les ARN en double brin est l’une des enzymes associée au mécanisme des ARN agissant comme antisens. Par la technique de RIP-Seq, de nombreuses cibles ARN ont été identifiées révélant de nouveaux mécanismes régulés par la RNase III. Ces mécanismes impliquent par exemple une maturation spécifique des ARNm conduisant à une stabilisation et une meilleure traductibilité, la maturation d’ARNm divergents ou convergents et possédant des régions non traduites chevauchantes, la régulation par les ARN antisens.
Quel est l’impact de la co-maturation des gènes divergents ou convergents sur la physiologie de la bactérie ? Y a t’il d’autres enzymes ou protéines associées à ces nouveaux mécanismes de régulation ?
Pour déterminer les protéines et/ou ARNm cibles impliqués dans la régulation post-transcriptionnelle, plusieurs approches dérivées de la littérature ont été adaptées chez S. aureus pour identifier des complexes régulateurs in vivo impliquant soit des ARNm ou des ARN non codant (ARNnc). Ces méthodes impliquent la séparation des complexes sur gradient de glycérol ou l’utilisation d’ARNm/ARNnc portant une étiquette pour séparer les complexes sur colonne d’affinité. L’identification des partenaires est ensuite réalisée par spectrométrie de masse (protéines) et RNA-seq (ARN cibles).
Ces approches permettent d’établir les réseaux de régulation. La fonction et le mécanisme d’action des co-facteurs des ARN sont actuellement étudiés.
